
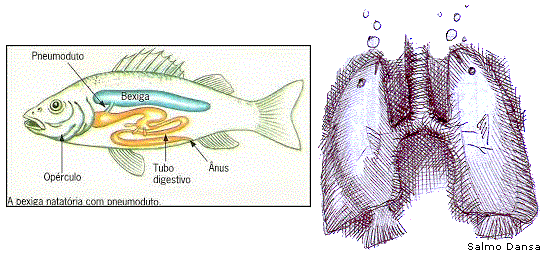
Bexiga natatória de peixes: um precursor do pulmão humano, como diz o evolucionismo?
Darwinistas frequentemente utilizam os peixes dipnoicos, ou seja, peixes ósseos portadores de bexiga-natatória como um exemplo do suposto “fato” de que esse órgão originalmente projetado para uma finalidade (flutuação) poderia ser convertido noutro órgão para uma finalidade completamente diferente (respiração) [1]. Portanto, sob a visão darwinista, o sistema respiratório humano nada mais é do que uma bexiga natatória avançada que foi otimizada por meio da seleção natural e mutações genéticas. A alegação é de que Darwin pôs um fim à questão ao mostrar a possibilidade de que as bexigas natatórias pudessem se transformar em pulmões em anfíbios (a propósito, não demonstrado por fósseis) evoluindo posteriormente para pulmões de répteis, de aves e de mamíferos (especialmente, os pulmões de humanos).
A estória começa da seguinte forma: peixes precisavam de um ambiente mais eficiente para atender o aumento da demanda de oxigênio. Para isso, eles inicialmente passaram a se concentrar perto da superfície da água (a propósito, estando expostos à predação). Então, a partir das brânquias a evolução deu origem à bexiga natatória, que depois supostamente teria resultado nos pulmões. Mas a estória não para por aí, podendo ser encontrada na maioria dos livros didáticos:
Perto do fim do Devoniano [supostamente mais de 250 milhões de anos atrás] uma das linhagens de peixes barbatanas-lobo conseguiu se mover para a terra. As barbatanas destes animais, já grossas e musculosas, foram talvez adaptadas para mover-se lentamente através de águas rasas e vegetação aquática espessas; seus antepassados já haviam começado há muito tempo atrás a contar com a respiração do ar, assim como muitos peixes que vivem em águas estagnadas hoje […] Quarenta milhões de anos após as incursões pioneiras de plantas, a invasão dos vertebrados de terra estava em curso. [2: p.559]
O naturalista britânico Charles Darwin observou e descreveu a suposta evolução das estruturas especializadas que participam da respiração dos peixes da seguinte forma:
dois órgãos distintos por vezes realizam simultaneamente a mesma função no mesmo indivíduo; para dar um exemplo, há peixes com guelras ou brânquias que respiram o ar dissolvido na água, ao mesmo tempo que respiram o ar livre nas suas bexigas-natatórias, sendo este último órgão munido de um ductus pneumaticus para o seu abastecimento, dividindo-se em partições vasculares. […] Nestes casos, um dos dois órgãos poder-se-ia modificar e aperfeiçoar facilmente, de maneira a realizar por si todo o trabalho, sendo auxiliado durante o processo de modificação pelo outro órgão; e então este outro órgão poderia ser modificado para outra finalidade completamente distinta, ou poderia ser completamente eliminado. [1: p.147]
Bela especulação! Porém, o que Darwin não sabia é que o oxigênio entraria na bexiga natatória de peixes pelo sangue em vez de uma traquéia [3]. Somente décadas mais tarde é que se descobriu que “o transporte ativo de oxigênio na bexiga natatória pela glândula gás é um transporte de oxigênio molecular” [4: p.521]. Portanto, tudo aponta para um sistema irredutivelmente complexo. Como bem observaram Brauner e colaboradores:
A transição da respiração aquática para a aérea está associada a mudanças fisiológicas dramáticas em relação à troca gasosa, regulação iônica, equilíbrio ácido-base e excreção de resíduos nitrogenados. [5: p.1433].
Após mais de um século, essa hipótese ainda hoje apresenta muitos obstáculos. Se não bastasse, como resolver a mais uma situação nitidamente irônica: o peixe iria sufocar se ele estivesse exposto ao ar durante longos períodos [3]. Isso porque a concentração de oxigênio dissolvido em água é de cerca de um trigésimo em relação ao ar [6: p.284]. Mas mesmo com este aumento de oxigênio, um peixe iria rapidamente se asfixiar se ele fosse levado para fora da água, embora estivesse em um ambiente com aumento de oxigênio. Brânquias são inoperáveis em ambientes secos, e líquidos destroem fisicamente alvéolos nos pulmões [6: p.284].
Mas, então, qual seria a evidência que Darwin possuía para afirmar a transmutação de um órgão noutro? Bem, nenhuma! Darwin alegou que todos os fisiologistas da época admitiam que
a bexiga-natatória é homóloga, ou ‘idealmente semelhante’ em posição e estrutura, aos pulmões dos animais vertebrados superiores. [1: p.148]
Para ele, então, a homologia seria suficiente para acreditar que a seleção natural converteu efetivamente uma bexiga-natatória num pulmão, ou num órgão exclusivamente usado para respirar. Porém, como afirma o biólogo molecular Dr. Michael Denton:
A base evolutiva da homologia é talvez ainda mais severamente danificada pela descoberta de que estruturas aparentemente homólogas são especificadas por diferentes genes em espécies diferentes. [7: p.149]
Como bem descreveu Darwin, os peixes dipnoicos (a Pirambóia, por exemplo) possuem um duplo sistema para a respiração. Além de guelras, eles possuem a bexiga natatória (pulmão), sendo essa característica a que tem levado os darwinistas a verem nele um vínculo entre os anfíbios e os peixes. Mas teria esses peixes realmente alguma ligação com os anfíbios que supostamente deram origem aos vertebrados terrestres? A resposta é não! Primeiro, porque os peixes dipnoicos fazem parte de um grupo totalmente separado [8: p.65]. Segundo, porque os ossos de seus crânios diferem dos ossos dos primeiros anfíbios fósseis encontrados [9: p.137]. E, por último, porque os próprios paleontologistas evolucionistas assumem que
a origem das principais características de tetrápodes [anfíbios] permaneceu obscura por falta de fósseis que documentem a sequência das mudanças evolutivas. [10: p.757]
Até pouco tempo atrás, por exemplo, era “fato” que os anfíbios evoluíram do peixe Celacanto, que possui nadadeiras lobadas (isto é, com projeções ósseas robustas) e uma bexiga natatória cheia de óleo. Porém, novas análises filogenéticas estão fazendo um verdadeiro estrago na classificação taxonômica evolutiva baseada em aspectos morfológicos. Atualmente, os peixes com bexigas natatórias têm sido considerados mais intimamente relacionados com os mamíferos do que com o próprio Celacanto [11]. Contudo, não podemos nos esquecer que peixes com bexiga natatória (pulmonados) têm tamanhos do genoma aproximado de 133 bilhões de pares de base enquanto que os seres humanos têm um tamanho de genoma de cerca de 3,5 bilhões de pares de bases [12]. Na melhor das hipóteses, baseado puramente em tamanho do genoma, poderíamos dizer que o ser humano compartilha uma similaridade de apenas 3% com os peixes pulmonados.
Um golpe ainda mais amargo para os evolucionistas (que não veremos em livros didáticos) é que o registro fóssil veio à tona forçar uma reversão de 180 graus na estória sobre a “bexiga natatória para pulmão”. A presença de pulmões em fósseis de placodermos (classe de peixes extintos) demonstra serem os pulmões muito mais “antigos” do que as bexigas natatórias, o que sugere que todos os peixes primitivos Jawed também já o teriam [13, 14]; então, seguindo esse raciocínio, os pulmões é que deveriam ter evoluído para bexigas natatórias, ao contrário do que seria esperado.
Mas o que a fisiologia dos peixes tem a nos dizer?
Os peixes dipnoicos possuem a bexiga natatória, um órgão projetado para que estas criaturas melhorem seu desempenho ao subirem e descerem na água. A bexiga natatória possui uma glândula que permite a expansão ou encolhimento desse órgão, alterando, portanto, o volume da entrada de gás em seu interior [15]. Os gases acumulados na bexiga natatória apresentam em sua composição Oxigênio, Gás carbônico e Nitrogênio [16]. Aliás, é devido a esses gases no interior da bexiga natatória que os peixes boiam, após mortos. A concentração desses gases varia de espécie para espécie.
Ademais, em algumas espécies de peixes, principalmente de água doce, o grau de funcionalidade desse projeto vai além: a bexiga encontra-se ligada com o labirinto do ouvido interno, onde o animal obtém um sentido preciso da pressão da água (e, portanto, da profundidade) [17]. Por esse motivo acredita-se que esta ligação também auxilia o sentido da audição dos peixes. Se não bastasse, alguns peixes fazem com que suas bexigas natatórias vibrem por meio do uso de músculos super-rápidos a fim de produzir e emitir sons, característica esta vantajosa para o acasalamento e geração de descendentes [18].
Outro ponto interessante diz respeito ao mecanismo de flutuação da bexiga natatória, o qual proporciona flutuabilidade neutra perfeita a um peixe e ele pode desse modo evitar gasto de energia para não afundar [16]. Há, entretanto, uma regra importante para que esse projeto se mantenha funcional e otimizado: o peixe deve permanecer a uma profundidade específica. Se nadar abaixo dessa profundidade, a bexiga será comprimida pelo aumento da pressão na água, a flutuabilidade diminuirá e o peixe precisará nadar ativamente para evitar que afunde mais. Também já se sabe que o fato de um determinado peixe possuir uma bexiga natatória está mais relacionado ao ambiente em que vive do que a sua posição taxonômica. Alguns peixes que vivem no fundo, por exemplo, são desprovidos de bexigas natatórias e parece razoável que a flutuabilidade neutra não lhe seja necessária.
Nós, proponentes do design, não contestamos a ideia de que bexigas natatórias e os pulmões se desenvolvem a partir dos mesmos tecidos básicos, embora a hipótese específica da “origem a partir” do divertículo do intestino anterior seja desafiada [19]. No modelo evolucionista, isso é interpretado em termos de ancestralidade comum; no modelo do design inteligente, em termos do mesmo projeto básico de construção, com variações criativas sobre o mesmo tema. Se dotado com brânquias, pulmões, ou uma combinação de ambos (a Pirambóia, por exemplo), todos os peixes, vivos ou extintos, parecem ser (ou terem sido) bem equipados para as exigências do seu modo de vida. Portanto, o chamado “fato” da evolução de pulmões de bexigas natatórias finalmente (e evidentemente) acaba por ser apenas mais um mito.
Fonte: Capítulo disponível no web book: Teoria do Design Inteligente
Referências:
[1] Darwin CR. The Origin of Species by means of Natural Selection. 6. Ed. London: John Murray, 1872, Capítulo 6. [Link]
[2] Starr C, Taggart R. Biology: The Unity and Diversity of Life. Belmont, CA: Wadsworth Publishing, 1987.
[3] Harrub B. The Breath of Life—Not a Product of Evolution. Reason & revelation 2006; 26(2):9-15. [Link]
[4] Wittenberg JB. The secretion of oxygen into the swimbladder of fish. J Gen Physiol. 1961 Jan;44:521-6.
[5] Brauner CJ, Matey V, Wilson JM, Bernier NJ, Val AL. Transition in organ function during the evolution of air-breathing; insights from Arapaima gigas, an obligate air-breathing teleost from the Amazon. J Exp Biol. 2004 Apr;207(Pt 9):1433-8.
[6] Maina JN. Structure, function and evolution of the gas exchangers: comparative perspectives. J Anat. 2002 Oct;201(4):281-304.
[7] Denton M. Evolution: A Theory in Crisis. Bethesda, MD: Adler & Adler, 1986.
[8] Ommanney FD. Os peixes. Trad. José Laurêncio de Melo. Série: Biblioteca da natureza life., Rio de Janeiro: Livraria José Olympio Editora, 1981.
[9] Attenborough D. Life on Earth: A Natural History. New York: Little, Brown & Co, 1981.
[10] Daeschler EB, Shubin NH, Jenkins FA Jr. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature. 2006 Apr 6;440(7085):757-63.
[11] Amemiya CT, et al. The African coelacanth genome provides insights into tetrapod evolution. Nature. 2013 Apr 18;496(7445):311-6.
[12] Leitch IJ. Genome sizes through the ages. Heredity (Edinb). 2007 Aug;99(2):121-2.
[13] Denison RH. A anatomia mole de Bothriolepis. J. Paleontol. 1941; 15:553-561.
[14] Romer AS, Parsons TS. The Vertebrate Body. 5. ed. Philadelphia: Saunders Co., 1978, p. 329.
[15] Setaro JF. Circulatory System. Microsoft Encarta, 1999.
[16] Schmidt-Nielsen K. Fisiologia Animal, Adaptação e Meio Ambiente. 5. Ed. Cambridge University Press, 2002, p. 609.
[17] Schulz-Mirbach T, Heß M, Metscher BD, Ladich F. A unique swim bladder-inner ear connection in a teleost fish revealed by a combined high-resolution microtomographic and three-dimensional histological study. BMC Biol. 2013 Jul 4;11:75.
[18] Mok HK, Parmentier E, Chiu KH, Tsai KE, Chiu PH, Fine ML. An Intermediate in the Evolution of Superfast Sonic Muscles. Front Zool. 2011 Nov 29;8(1):31.
[19] Brown E, James K. The lung primordium an outpouching from the foregut! Evidence-based dogma or myth? J Pediatr Surg. 2009 Mar;44(3):607-15.


Essa foi a maior besteira que eu já li em toda a minha vida. Estou traumatizada. Não sei sequer o que dizer.