

O arquiteto da vida: A complexidade fantástica do olho
Que o olho humano é uma maravilha tecnológica, ninguém discorda. Tanto é assim, que os engenheiros tentam há anos imitar a funcionalidade do nosso órgão da visão. Graças a esse esforço, a admiração deles pelo olho tem crescido cada vez mais. Os cientistas se deram conta de que nenhum chip de computador consegue fazer o que a retina faz. Se conseguisse, esse dispositivo teria que ser um milhão de vezes maior que a retina e pesaria mais de 40 kg! (A retina pesa menos de um grama e tem espessura de 0,01 milímetro)
E o consumo de energia?
Nossa retina consome apenas 0,0001 watt, enquanto a retina artificial requereria muito mais energia: 300 watts – além de um sistema de refrigeração.
Com tudo isso, teríamos finalmente a visão humana reproduzida artificialmente?
Ainda não. Se fosse construído um chip desses, ele seria capaz de lidar com uma área quadrada de apenas duas mil unidades de visão (pixels), enquanto a visão humana tem capacidade cinco vezes maior. Se tal chip existisse, teria equivalente a um milhão de transístores, enquanto a retina humana equivale a 25 bilhões deles!
Nota: Charles Darwin admitiu em “A origem das espécies“, de 1859, que parecia absurdo pensar que a estrutura ocular se desenvolveu por seleção natural. Os criacionistas e defensores do Design Inteligente defendem que o olho humano (assim como os de quaisquer vertebrados ou invertebrados) é fruto de um projeto complexo e inteligente em todos aspectos. Mas, contrariando Darwin e os criacionistas, os sucessores modernos de Darwin afirmam que o olho nada tem de projeto inteligente e que na verdade possui “falhas ou cicatrizes da evolução” que segundo eles, são evidências para descartar um Design Inteligente ou a criação por parte de Deus.
Quais seriam essas “falhas” ou “cicatrizes da evolução”?
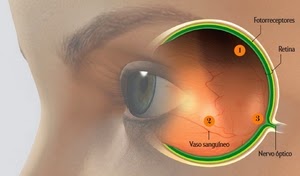 De acordo com os evolucionistas, o olho dos vertebrados, longe de ser concebido de forma inteligente, contém inúmeros defeitos que degradam a qualidade da imagem:
De acordo com os evolucionistas, o olho dos vertebrados, longe de ser concebido de forma inteligente, contém inúmeros defeitos que degradam a qualidade da imagem:
1ª Cicatriz: Retina invertida, que força a luz a atravessar corpos celulares e fibras nervosas antes de atingir os fotorreceptores;
2ª Cicatriz: vasos sanguíneos que se espalham pela superfície interna da retina, provocando sombras indesejadas;
3ª Cicatriz: fibras nervosas que se juntam, projetam-se numa abertura única na retina e viram o nervo óptico, criando um ponto cego.
Então o olho humano não foi projetado inteligentemente porque possui essas “cicatrizes evolutivas”?
Será que elas seriam realmente falhas (cicatrizes) e por isso elas são evidências de que não existe nenhuma inteligência superior (Design Inteligente) por trás da complexidade do olho?
Em primeiro lugar, precisamos revisar alguns pontos sobre a anatomia ocular:
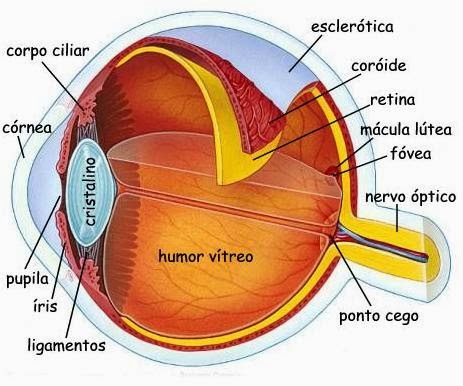 A luz entra no olho humano pela transparente córnea, a janela frontal ocular, que age como uma poderosa lente convexa. Após passa pela pupila (a abertura no diafragma da íris) a luz e posteriormente refratada pela cristalino.
A luz entra no olho humano pela transparente córnea, a janela frontal ocular, que age como uma poderosa lente convexa. Após passa pela pupila (a abertura no diafragma da íris) a luz e posteriormente refratada pela cristalino.
Uma imagem do ambiente externo é então focalizada na retina que transforma a luz em sinais nervosos e é a mais profunda (em relação ao centro do globo ocular) das três “túnicas” do segmentos posterior do olho.
As outras duas são a altamente fibrosa esclerótica (branco dos olhos) encontrada na parte mais externa continuando desde a córnea e a coroide, uma camada altamente pigmentada e irrigada que se encontra entre as outras duas.
A retina consiste de 10 camadas, das quais a mais externa é a escura camada do Epitélio Pigmentar da Retina (RPE em inglês) que devido ao pigmento melanina, é opaca a luz. Suas células tem finas projeções capilares em sua superfície interna chamada de “microvilosidades” que permeiam a região e recobrem as pontas dos fotorreceptores dos segmentos externos.
Daí porque existir um potencial ponto de fissão entre o RPE e os fotorreceptores que se torna evidente quando a retina se separe do RPE, como durante o resultado de uma lesão, uma condição conhecida como “deslocamento de retina”. Cada receptor, seja um bastonete ou cone, consiste de um segmento interior e um exterior, o primeiro contendo organelas para a fabricação dos pigmentos fotossensíveis presente no último. A camada de cones e bastonetes e todas as 8 camadas internas constituem o que é conhecido como “retina neurosensorial” que é virtualmente transparente à luz.
Por meio de inúmeras conexões nervosas complexas dentro da retina neurosensorial, impulsos elétricos gerados pela luz ao atingirem os fotorreceptores são processados e transmitidos a camada de fibras nervosas da retina, de onde seguem pelo nervo ótico até o cérebro.
Em muitas espécies que necessitam de visão em ambientes de baixíssima iluminação, uma camada de material cristalino reflexivo, o tapetum (tapete em latim) é incorporado ao RPE ou a coroide. Agindo como um espelho, o tapetum reflete a luz que passa entre os fotorreceptores, aumentando a quantidade de luz que atinge os receptores.
O epitélio pigmentar da retina
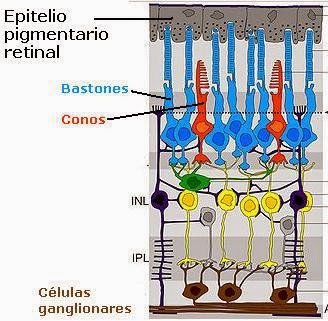 Fundamental para o entendimento da retina invertida é o papel crucial do RPE. Muitas de suas essenciais funções ainda não são bem conhecidas. Cada célula do RPE está em contato intimo com as pontas dos 20 ou mais segmentos fotorreceptores exteriores cujo número chega a 130 milhões. Sem o RPE os receptores e o resto da retina neurosensorial não podem funcionar plenamente e acabariam se atrofiando.
Fundamental para o entendimento da retina invertida é o papel crucial do RPE. Muitas de suas essenciais funções ainda não são bem conhecidas. Cada célula do RPE está em contato intimo com as pontas dos 20 ou mais segmentos fotorreceptores exteriores cujo número chega a 130 milhões. Sem o RPE os receptores e o resto da retina neurosensorial não podem funcionar plenamente e acabariam se atrofiando.
O segmento externo de um receptor consiste de uma pilha de discos contendo pigmentos fotossensíveis. Estes discos são constantemente formados no segmento interno de onde eles seguem sucessivamente dos segmentos externos em direção ao RPE que recicla seus componentes químicos pelo processo de “fagocitose” (do grego:φάγω (phagō) = comer).
O RPE armazena vitamina A, um precursor dos pigmentos fotossensíveis e em seguida participa de suas regenerações. Existem quatro fotopigmentos que se desbotam sob exposição à luz: rodopsina (encontrada em bastonetes, para visão noturna) e mais três tipos um para cada tipo de cone (um cone para cada cor primária). Ele sintetiza glicosaminoglicanas para o interfotorreceptor matriz, ou seja, o material interno que separa os receptores.
Além de oxigênio, o RPE seletivamente transporta nutrientes da coroide para suprir as três camadas externas da retina e remove os produtos residuais metabólicos dos receptores para serem varridos para circulação coroidal. Por meio de seletivo bombeamento de metabólitos e a presença de estreitas junções intercelulares, o RPE age como uma barreira, prevenindo o acesso de resíduos químicos maiores ou nocivos ao tecido retinal, com isso contribui com a constante manutenção de um ambiente retinal estável e otimizado.
O RPE possui complexos mecanismos para lidar com molécula tóxicas e radicais livres produzidos pela ação da luz. Enzimas especializadas como superóxido dismutase, catalases e peroxidases estão presentes para catalizar a quebra de moléculas potencialmente danosas como superóxido e o peróxido de hidrogênio. Antioxidantes como o atocoferol (vitamina E) e ácido ascórbico (vitamina C) estão presentes para reduzir os danos da oxidação.
Nossos receptores continuamente sintetizam novos discos com seus respectivos pigmentos fotossensíveis reciclando materiais dos discos usados que foram antes digeridos pelo RPE. Isto levanta a questão: Porque existe esse complicadíssimo processo?
A resposta pode estar no fato desse processo ser um exemplo de renovação biológica, pelo qual tecidos expostos a substâncias tóxicas, radiação, trauma físico, etc, podem sobreviver. Sem auto-renovação, tecidos como a pele, o revestimento do intestino, células sanguíneas e todo o resto iriam rapidamente acumular defeitos fatais. Do mesmo jeito, pela continua substituição dos discos os fotorreceptores superam o incessante processo de desintegração acelerado por agentes tóxicos, em particular, por luzes de comprimento de ondas curtas. Mas, até que o tecido “aprendesse” a se auto-renovar biologicamente, como poderia evoluir se constantemente estaria se desintegrando? [nosso questionamento]
O dissipador térmico coroidal
Tem sido observado que o dano aos receptores de modelos experimentais está fortemente relacionado com a temperatura, e outros estudos confirmam que calor potencializa lesões fotoquímicas. Qualquer sistema projetado para resistir ao último deve também proteger contra o primeiro. Em 1980, um artigo foi publicado explicando pela primeira vez algo já conhecido sobre a coroide. Isto é, sua altíssima taxa de fluxo sanguíneo que excede em muito a demanda nutricional da retina, apesar desta ser altamente ativa, metabolicamente, como já indicado.
Os vasos capilares coroidais (coriocapilares) formam um rico emaranhado na parte externa do RPE, predominantemente em sua área central, e separado deste apenas por uma fina membrana (membrana de Bruch). A absorção de luz excessiva pelo RPE produz calor na parte externa da retina que deve ser dissipado, afim de evitar danos pelo excesso de aquecimento ao delicado e complexo aparato biológico, e também a suas circunvizinhanças.
Os autores deste estudo contundentemente demonstram uma função essencial da coroide com seu fluxo torrencial sanguíneo (em termos locais) e sua proximidade ao RPE, ao funcionar como dissipador de calor e resfriador. Ainda mais fascinante são os resultados de estudos posteriores pelos mesmos autores indicando que existem reflexos nervosos centrais (pelo cérebro) mediados pela luz que regulam o fluxo sanguíneo coroidal, aumentando o fluxo sanguíneo conforme a iluminação aumenta. Ambos RPE e coroide tem de estar localizados na parte externa da retina neurosensorial; disso podemos concluir que existem sólidas razões para a configuração invertida da retina de humanos e outros vertebrados. (Parver, L.M., Temperature modulating action of choroidal blood flow, Eye 5:181, 1991.); (Parver, L.M., Temperature modulating action of choroidal blood flow, Eye 5:181, 1991.)
A fóvea
Embora a retina neurosensorial seja virtualmente transparente excetuando-se em seus finíssimos vasos sanguíneos, existe um refinamento adicional de sua estrutura em sua região central chamada de mácula lútea. A retina e o córtex cerebral ocipital (chamado de córtex visual), para o qual a retina transmite informações visuais, é tão organizado que o VA é máximo no eixo visual. O eixo visual passa pela fovéola que forma o “piso” de uma fenda circular com uma parede declinada, chamado de fóvea (do latim= fenda, fossa) no centro da mácula. Longe da fóvea o VA diminui progressivamente em direção a periferia da retina. Daí os fotorreceptores de cor- os cones que detectam vermelho, verde e possivelmente azul- tem a maior densidade, de 150.000 por mm² na fovéola, que mede apenas 300-330 µm (micrometro) de extensão.
O pigmento xantofila
O sistema ótico do olho humano é projetado de tal maneira que a luz ambiente tende a atingir com toda intensidade a área macular da retina, e com muito menos intensidade a periferia desta. É significante que não apenas a quantidade de melanina é mais abundante na região macular devido a suas células RPE serem maiores e mais numerosas por milimetro quadrado do que em qualquer outro lugar, mas também no centro da retina encontra-se o pigmento xantofila (Grego: ξάνθος- xanthos, amarelo). Nesta região, a xantofila permeia todas as camadas da retina neurosensorial entre suas duas membranas e é concentrada nas células retinais, ambos os neurônios e células dos tecido que os sustenta. Atenção tem sido dada recentemente a presença de uma coleção de tecidos celulares de suporte chamada de tecidos de Muller que se encontram na superfície da fóvea e formam um cone cuja ponta pluga na depressão foveolar.
Xantofila é um carotenoide quimicamente ligado a vitamina A, cujo ápice do espectro de absorção chega a 460 nm e varia de 480 nm a 390 nm. Ela ajuda a proteger a retina neurosensorial ao absorver maior parte das ondas mais curtas da luz visível, ou seja, o espectro azul e violeta, potencialmente mais danosos.
O ponto cego
Devido a disposição invertida da retina, os axônios (fibras nervosas) que transmitem dados ao cérebro passam por baixo da superfície interna da retina e convergem em um pequeno ponto que é a “cabeça” do nervo ótico, daí, saindo para o cérebro. A cabeça do nervo ótico não possui receptores de luz, sendo portanto cega, causando um pequeno ponto cego no campo de visão. Sem surpresa, evolucionistas criticaram isso:
“Nosso ponto cego da retina raramente causa alguma dificuldade, mas raramente não quer dizer nunca. Quando por um momento cubro um olho para repelir um inseto, um evento importante pode estar focalizado no ponto cego do outro.” (Williams, G.C., Natural Selection: Domains, Levels and Challenges, Oxford University Press, Oxford, pp. 72–73, 1992.)
Além disso, o problema tem de ser visto em perspectiva: o ponto cego encontra-se a 15° do centro do eixo visual (a 3.7 mm da fovéola) e é minúsculo em relação ao campo de visão do olho, ocupando menos de 0.25% deste. Como mencionado acima, quanto mais longe da fovéola um ponto na retina está, menor será seu VA e sua sensibilidade.
A retina ao redor da cabeça do nervo ótico, possui um VA apenas 15% igual ao da fovéola. Podemos inferir com segurança que o risco em teoria, previsto por Williams decorrente do ponto cego em uma pessoa com apenas um olho é insignificante; e, em concordância com isso, é considerado seguro para uma pessoa de um olho só dirigir um carro para fins não profissionais.
Como os dois campos visuais se sobrepõem em grande parte, o ponto cego de um olho é coberto pelo campo visual do outro. É verdade que a oclusão ou perda de um dos olhos é uma desvantagem, mas isso não é por causa do ponto cego do olho que notamos pelas razões expostas acima.
Olhos dos invertebrados
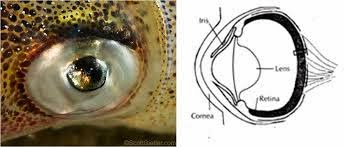 Existem algumas alegações de que a retina não-invertida de cefalópodes, tais como lulas e polvos, são mais eficientes do que a retina invertida encontrada em vertebrados. Mas isso pressupõe que a retina invertida é ineficiente, mas já vimos que não é o caso.
Existem algumas alegações de que a retina não-invertida de cefalópodes, tais como lulas e polvos, são mais eficientes do que a retina invertida encontrada em vertebrados. Mas isso pressupõe que a retina invertida é ineficiente, mas já vimos que não é o caso.
Além disso, nunca foi demonstrado que os cefalópodes realmente enxergam melhor. Pelo contrário, seus olhos apenas “se aproximam a alguns dos olhos de vertebrados inferiores em termos de eficiência” e são provavelmente cegos de cores (só enxergam preto e branco). Além disso, a retina de cefalópodes, além de ser “convertida”, é muito mais simples do que a retina “invertida” de vertebrados:
“A estrutura da retina [de cefalópodes] é muito mais simples do que do olho dos vertebrados, com apenas dois componentes neurais, células receptoras e fibras eferentes.” (Budelmann, B.U., Cephalopod sense organs, nerves and brain, 1994. In Pörtner, H.O., O’Dor, R.J. and Macmillan, D.L., ed., Physiology of cephalopod molluscs: lifestyle and performance adaptations, Gordon and Breach, Basel, Switzerland, p. 15, 1994.)
É uma estrutura ondulante com “longas células fotorreceptoras cilíndricas com omatídeos consistindo de microvilosidades”, portanto os olhos de moluscos tem sido descritos como um “olho composto com lentes simples”.
Finalmente, eles vivem em regiões com muito menos intensidade de luz do que a maioria dos vertebrados, o que contribui para demonstrar que olhos de cefalópodes não precisam ser tão complexos como é costumeiramente afirmado.
Conclusão
A luz em vários comprimentos de onda é capaz de efeitos muito nocivos para a maquinaria biológica. A retina, além de ser um processador e transdutor de imagem extremamente sofisticado, é claramente projetado para suportar os efeitos tóxicos e de aquecimento de luz. O olho está bem equipado para proteger a retina contra a radiação que normalmente encontramos na vida cotidiana. Além da quase completa exclusão de radiação ultravioleta pela córnea e a lente em conjunto, a própria retina é dotada com um certo número de mecanismos adicionais para proteger contra tais danos:
O epitélio do pigmento da retina produz substâncias que combatem o produto químico prejudicial dos subprodutos da radiação de luz e desempenha um papel essencial para sustentar os fotorreceptores. Isso inclui a reciclagem e metabolização dos seus produtos, assim renovando-as em face de desgaste contínuo ao bombardeio de luz.
A área central da retina é permeada com pigmento xantofila que filtra e absorve de curto comprimento de onda da luz visível. Os fotorreceptores, portanto, precisam estar em contato íntimo com o epitélio pigmentar da retina, que é opaco. O epitélio do pigmento da retina, por sua vez, necessita de estar em contato íntimo com a coroide (também opaca) para satisfazer as suas necessidades nutricionais e para prevenir (por meio do efeito do dissipador de calor do seu fluxo de sangue massiva) o sobreaquecimento da retina quando focada na luz.
Se a retina não fosse invertida (como é) e os fotorreceptores apontassem em direção a luz, e suas ligações nervosas em direção ao cérebro (configuração convertida) como propõem evolucionistas como Richard Dawkins (Dawkins, R., The Blind Watchmaker: Why the evidence of evolution reveals a universe without design. W.W. Norton and Company, New York, p. 93, 1986.), suas duas camadas opacas teriam de ser interpostas no trajeto da luz para os fotorreceptores, o que iria deixá-los na escuridão.
Assim, é sugerido que a necessidade de proteção contra os danos induzidos pela luz, que uma retina convertida no nosso ambiente natural não proporcionaria com o mesmo grau, é um dos principais, se não a principal razão para a existência da configuração invertida da retina.
Ainda que possa parecer, à primeira vista, que essas “cicatrizes” sejam desvantagens e são ineficientes como muitos evolucionistas afirmam, na realidade, como vimos, é exatamente o oposto.
E você, o que acha?
Será que a complexidade fantástica do olho é resultado de um processo acidental do acaso e do tempo? Ou é um projeto de criação elaborado pelo Arquiteto da vida?
Referências:
Darwinismo – Os sistemas de visão estão de acordo com a teoria da evolução?
Wol.JW – Teve um Projeto? A retina invertida
Criacionismo – O que é preciso para construir um olho?
Creation.com – Is our ‘inverted’ retina really ‘bad design’?
Imagens fonte: Reprodução Google



ótima explicação
amei abrigada